The Influence of Foot-Strike Technique on the Neuromechanical Function of the Foot
Abstract and Introduction
Abstract
Purpose: The aim of this study was to investigate the influence of foot-strike technique on longitudinal arch mechanics and intrinsic foot muscle function during running.
Methods: Thirteen healthy participants ran barefoot on a force-instrumented treadmill at 2.8 ms−1 with a forefoot (FFS) and rearfoot (RFS; habitual) running technique, whereas kinetic, kinematic, and electromyographic data from the intrinsic foot muscles were collected simultaneously. The longitudinal arch was modeled as a single "midfoot" joint representing motion of the rearfoot (calcaneus) relative to the forefoot (metatarsals). An inverse dynamic analysis was performed to estimate joint moments generated about the midfoot, as well as mechanical work and power.
Results: The midfoot was more plantar flexed (higher arch) at foot contact when running with a forefoot running technique (RFS 0.2° ± 1.8° vs FFS 6.9° ± 3.0°, effect size (ES) = 2.7); however, there was no difference in peak midfoot dorsiflexion in stance (RFS −11.6° ± 3.0° vs FFS −11.4° ± 3.4°, ES = 0.63). When running with a forefoot technique, participants generated greater moments about the midfoot (27% increase, ES = 1.1) and performed more negative work (240% increase, ES = 2.2) and positive work (42% increase, ES = 1.1) about the midfoot. Average stance-phase muscle activation was greater for flexor digitorum brevis (20% increase, ES = 0.56) and abductor hallucis (17% increase, ES = 0.63) when running with a forefoot technique.
Conclusions: Forefoot running increases loading about the longitudinal arch and also increases the mechanical work performed by the intrinsic foot muscles. These findings have substantial implications in terms of injury prevention and management for runners who transition from a rearfoot to a forefoot running technique.
Introduction
The orientation of a runner's foot at the moment of foot-ground impact is often used as a method to classify running technique.[1–4] Foot-strike patterns are generally classified into three groups: 1) runners who land in a heel-first manner (rearfoot strike), 2) runners who land on the forefoot (forefoot strike (FFS)), and 3) runners who land with a simultaneous heel and forefoot contact (midfoot strike).[2,5] Although the majority (>75%) of distance runners adopt a rearfoot running technique,[6,7] it has been suggested that a forefoot running technique is mechanically advantageous, because it affords the runner greater opportunity to recycle the energy associated with foot–ground impact via elastic stretch and recoil of the tendons and ligaments of the ankle and foot.[4,8,9] Consequently, the popularity of running "retraining" programs that promote a forefoot landing pattern to reduce injury risk and improve running performance has surged.[5,10,11]
The longitudinal arch (LA) of the human foot behaves in a springlike manner when running, temporarily storing and then subsequently returning a considerable portion of the mechanical energy required for each stride.[12–14] Compression (lowering and lengthening) of the LA during the first half of stance phase allows mechanical energy to be temporarily stored within the stretched ligaments, muscles, and tendons that span this structure.[12,14,15]This energy is subsequently returned in late stance, because the resultant ground reaction force (GRF) declines and the stretched elastic structures shorten to allow the LA to recoil (rise and shorten).[4,12,15]
Recently, studies have explored the hypothesis that a forefoot running technique enhances foot-spring function by allowing a greater proportion of mechanical energy to be recycled during each stride.[4,14,16] Consistent with the hypothesis of enhanced foot-spring function, these studies have all reported an increase in LA compliance with a forefoot technique compared with a rearfoot technique when running at matched velocities. However, the increased compliance seems to be due to a higher LA at foot contact, rather than an increase in peak LA compression in midstance.[4,14,16] A key finding in each of these studies was that peak LA compression and model-derived plantar aponeurosis strains were similar, regardless of foot-strike technique.[4,14,16] Given that peak strain primarily determines the magnitude of stored elastic energy, this finding suggests no energetic benefits to a forefoot running technique. The studies by McDonald et al.[16] and Wager and Challis[14] provide valuable insight into the biomechanics of the LA when running with rearfoot and forefoot running techniques. However, these studies have a methodological limitation in their assumption that the foot is a passive structure with no active muscular control of its biomechanical function.
Forefoot running is characterized by the center of pressure (COP) being located further anterior in the foot and higher peak GRF.[2,5,17,18] Thus, in a similar manner to the ankle joint,[19] an increase in the length of the GRF moment arm, combined with the higher peak GRF, will likely increase loading of the LA when running with a forefoot technique. Given that the intrinsic foot muscles have similar anatomical pathways to the plantar aponeurosis[20–22] and have the capacity to alter the stiffness of the LA,[15,23] it is highly likely that these muscles also contribute to the observed alterations in LA mechanics between foot-strike techniques. An increase in activation of the intrinsic foot muscles when running with a forefoot technique could potentially reduce excessive LA compression and reduce plantar aponeurosis strain, providing an explanation for the lack of difference in plantar aponeurosis strain observed in previous studies.[14,16]Furthermore, these muscles are also known to display preparatory activation before foot contact;[15,24] thus, it is conceivable that increased activation of these muscles in late swing may also explain the increased LA height at foot contact when running with a forefoot technique.
It is currently unknown if the intrinsic foot muscles contribute to the regulation of LA motion when switching from a rearfoot to a forefoot running technique. It is also unknown if loading of the LA changes when converting to forefoot running technique. Therefore, the aim of this study was to test the hypothesis that a forefoot running technique results in higher magnitudes of activation in the intrinsic foot muscles during the stance and swing phases while running, because of the higher mechanical loading occurring about the LA.
Methods
Participants
Thirteen healthy participants (five women and eight men; age, 24 ± 6 yr; height, 172 ± 8 cm; mass, 72 ± 12 kg) with no diagnosed history of lower limb injury in the previous 6 months or known neurological impairment volunteered to participate in the study. All participants were habitually shod recreational runners, who classified themselves as non–forefoot strikers. Written informed consent was obtained from each subject. The study protocol was approved by the institutional human research ethics committee and conducted in accordance with the Declaration of Helsinki.
Experimental Protocol
After a 3-min warm-up period and familiarization procedure, participants ran barefoot on a force-instrumented treadmill (AMTI, force-sensing tandem treadmill; Watertown, MA) at 2.8 m·s−1 with their habitual foot-strike (HFS) pattern and an FFS pattern, in a counterbalanced order. During the familiarization session, foot-strike pattern was confirmed by the investigator using visual analysis. None of the participants displayed a habitual forefoot running technique. Before commencement of each experimental condition, participants were given as much time as necessary to familiarize themselves with running on the treadmill with an FFS pattern. Participants were considered familiarized with the forefoot running technique once they were able to perform 20 s of consecutive FFS at the experimental speed and reported to be comfortable running with this technique. Foot-strike pattern during the familiarization period was confirmed visually by the investigator (L.K.). Kinetic, kinematic, and electromyographic (EMG) data were collected simultaneously for approximately 15–20 strides (toe-off to ipsilateral toe-off) across a 15-s data collection period for each running condition (HFS and FFS).
Data Acquisition
Kinematic and kinetic measurements. Motion data were captured at 200 Hz using an eight-camera three-dimensional optoelectronic motion capture system (Oqus; Qualysis, Gothenburg, Sweden). GRF and EMG data were synchronously captured with the motion data at 4000 Hz using a 14-bit analog to digital converter and the Qualisys Track Management software (Qualisys).
Three-dimensional motion of the foot and shank was modeled from motion capture data during each running trial. To quantify motions of the foot segment and the LA,[24,25] retroreflective markers (9.0-mm diameter) were secured on the skin of the right foot overlying the medial and lateral malleoli, posterior calcaneus, navicular tuberosity, and head of the first and fifth metatarsals. Additional markers were applied to the medial and lateral femoral condyles, and a rigid cluster of four markers was placed on the anterolateral aspect of the shank. During a standing calibration trial, markers located on the segment end points were used to generate a two-segment model of the shank and foot. A virtual marker (med-knee) corresponding to the position of the medial marker was also generated to reduce the influence of soft tissue artifact on skin-mounted markers located at the knee. After the calibration trial, the medial and lateral knee markers were removed and the motion of the shank and med-knee virtual marker was tracked using the rigid marker cluster located on the shank. Markers were adhered with double-sided adhesive and further secured with cohesive bandage, allowing for secure positioning for both running conditions. Markers were not removed between experimental trials.
EMG. Identification of the abductor hallucis (AH) and flexor digitorum brevis (FDB) muscles in the right foot of each subject was conducted using real-time B-mode ultrasound imaging (10-MHz linear array, SonixTouch; Ultrasonix, BC, Canada). Subsequently, bipolar fine-wire electrodes (0.051-mm stainless steel, Teflon-coated wires; Chalgren, Gilroy, CA) with a detection length of 4 mm and an interelectrode distance of 4 mm were inserted using delivery needles (0.5 mm × 50 mm) into the muscle tissue of AH and FDB under ultrasound guidance, in accordance with previously described methods.[26] Sterile techniques were used for the insertion of all wires. Surface EMG data were additionally collected from medial gastrocnemius (MG) and soleus (SOL) from the right leg of all participants using Ag–AgCl electrodes with a diameter of 10 mm and an interelectrode distance of 20 mm (Tyco Healthcare Group, Neustadt, Germany). A surface reference electrode, of the same type and size, was placed over the right fibula head. Before electrode placement, the areas of the leg corresponding to the electrode placement sites were shaved, lightly abraded, and cleaned with isopropyl alcohol. All EMG signals were amplified 1000 times and hardware filtered with a bandwidth of 30 to 1000 Hz (MA300; Motion Labs, Baton Rouge, LA). To minimize movement artifacts, the fine-wire electrodes, surface electrodes, connectors, cabling, and preamplifiers were secured with cohesive bandage around the foot and shank.
Before data collection, each participant was asked to perform foot maneuvers known to activate each of the muscles separately. Activation of the AH muscle was assessed by asking the participant to abduct and flex the great toe about the metatarsophalangeal joint while avoiding flexion of the interphalangeal joint. The FDB activation was assessed by asking the participant to flex toes 2–4 about the metatarsophalangeal joints while minimizing flexion of the distal interphalangeal joints.[26,27] When predicted EMG patterns could be detected, it was concluded that the fine-wire electrodes were in the correct location. If not, the electrodes were withdrawn by approximately 1 mm until appropriate activation patterns could be detected and possible crosstalk excluded. To confirm signal integrity and quality between trials, the EMG signals were retested after each trial using the same foot maneuvers applied previously. A Velcro strap was secured around the participant's waist, which enabled the EMG amplifier box to be secured to the subject without interfering with their gait. A lightweight optical cable connected the amplifier box to the analog to digital converter that was positioned well away from the subject and treadmill.
Data Analysis
Marker trajectories and GRF data were exported to Matlab (The Mathworks Inc., Natick, MA), whereas EMG data files were exported to Visual3D (C-motion Inc., Germantown, MD) for postprocessing and analysis.
Ground reaction forces. GRF data were filtered using a 35-Hz recursive, low-pass second-order Butterworth filter. A vertical GRF threshold was set to define each toe-off as occurring when vertical GRF fell below 50 N, whereas foot contact was defined as occurring when vertical force rose above 50 N. The magnitude of the peak vertical and anteroposterior components of the GRF was calculated and normalized to body weight (BW) for each participant. Peak propulsive force was defined as the peak positive value of the anteroposterior component of the GRF.
Stride temporal characteristics. Swing phase was defined as the period from right toe-off to right foot contact, whereas stance phase was defined as occurring between right foot contact and right toe-off. One stride cycle was considered as right toe-off to the subsequent right toe-off on the basis of GRF data. Stride duration and ground contact times were calculated for both conditions, for all participants.
Joint kinematics. Before calculation of joint kinematics, marker trajectories were low-pass filtered using a recursive, second-order Butterworth filter at 20 Hz. Initially, an assumed rigid segment was created for the foot.[24,25] All marker data were transformed to the foot segment coordinate system, allowing all sagittal plane angles to be represented relative to the orientation of the z-axis of the foot. Thus, all sagittal plane rotations represent flexion–extension of their respective joints. Ankle angle was defined as the planar angle created by the bisection of a vector projecting from the medial malleolus to the med-knee virtual marker, with another vector projecting from the head of the first metatarsal to the medial malleolus, projected onto the sagittal plane of the foot. Ankle angle was offset to the ankle angle during quiet bipedal stance, calculated during the standing calibration trial. Foot-strike angle was defined as the sagittal plane ankle angle at foot contact. Minimum ankle angle was defined as the minimum angle during stance phase, representing peak ankle dorsiflexion. Ankle angle at toe-off was also calculated.
To gain a deeper understanding of the mechanical behavior of the foot, we also modeled the foot as a two-segment structure with rearfoot and midfoot joints. The rearfoot joint angle was defined as the internal angle created by the bisection of a vector projecting from the med-knee virtual marker to the medial malleolus, with another vector projecting from the navicular marker to the medial malleolus marker, projected onto the sagittal plane of the foot (see Figure, Supplemental Digital Content 1, Depiction of sagittal plane angular rotations about the ankle (top), rearfoot (middle) and midfoot (bottom) joints, http://links.lww.com/MSS/B37. Angles in the text are relative to the respective joint angle in quiet stance, with 0° representing the joint angle in quiet stance). Positive rearfoot joint angles indicate plantar flexion of the rearfoot relative to the shank. Thus, the rearfoot angle is representative of the ankle joint in our multisegment foot model.
The LA is a complex structure that comprised a series of articulations made by a number of different bones.[28,29] Given the difficulties in quantifying the rotations about a series of small joints, located in such close proximity, the LA is often modeled as a single functional "midfoot" joint, representing the generalized angular motion of this structure.[15,24,25,30,31] Therefore, the biomechanical function of the LA during running was represented by a midfoot joint, defined as the internal angle created by the bisection of a vector projecting from the medial malleolus marker to the navicular marker (rearfoot segment) and another vector projecting from the head of the first metatarsal to the navicular marker, projected onto the sagittal plane of the foot[25] (see Figure, Supplemental Digital Content 1, Depiction of sagittal plane angular rotations about the ankle (top), rearfoot (middle) and midfoot (bottom) joints, http://links.lww.com/MSS/B37. Angles in the text are relative to the respective joint angle in quiet stance, with 0° representing the joint angle in quiet stance). A decrease in midfoot angle signifies dorsiflexion, or LA compression (upward rotation of the distal end of the forefoot segment) relative to the rearfoot. The rearfoot and midfoot angles were offset to the corresponding angles calculated during quiet bipedal stance, so that 0° represents the angle in quiet bipedal stance. Angle at foot contact, minimum stance angle (peak dorsiflexion), and angle at toe-off were calculated separately for the rearfoot and midfoot joints, for all participants.
Joint kinetics. An inverse dynamic analysis was performed using custom written software (Matlab; The Mathworks Inc.), allowing for estimation of net joint moments about the ankle, rearfoot, and midfoot during stance phase. Unfiltered marker trajectories and GRF and COP data were expressed relative to the foot coordinate system. Subsequently, proximal segment forces were resolved from the ground up using a link segment model.[32]The midfoot segment was defined as the vector projecting from the navicular marker to the marker located on the head of the first metatarsal, projected onto the sagittal plane of the foot. The rearfoot segment was defined as the vector projecting from the medial malleolus marker to the navicular marker, projected onto the sagittal plane of the foot. A traditional rigid foot segment was also defined to estimate ankle joint moments, enabling comparison of multisegment foot kinetics with traditional calculations on the basis of the assumption of a rigid foot segment. Segment masses and inertial properties were incorporated from previous literature,[31,33] with the center of mass location for each segment being defined as 50% of the distance between the segment end points.[31] Each joint was modeled as a pin joint with rotation occurring about the joint created by the bisection of the two vectors. Each joint axis was oriented parallel to the z-axis of the foot coordinate system. After estimation of joint moments, joint powers were calculated for the stance phase by multiplying the net joint moment by the angular velocity of each corresponding joint.[32] During early stance in a rearfoot running technique, GRF is applied to the rearfoot and forefoot segments simultaneously when the foot is flat on the ground. To overcome the difficulty in determining the relative percentage of GRF that is acting on the forefoot and rearfoot segments during early stance, joint moments for the rearfoot and midfoot were only calculated during stance phase after the COP had progressed distal to the axis of the midfoot joint, minimizing any potential errors associated with segment force assignments.[34] Subsequently, joint moments and joint powers for the ankle, rearfoot, and midfoot were filtered using a 20-Hz recursive, second-order low-pass Butterworth filter.[35] Net negative and positive work performed by each joint during stance phase was calculated by applying a trapezoidal integration of joint power curves across the stance phase. Quasi-midfoot stiffness was quantified by dividing the change in midfoot joint moment by the change in midfoot angle during stance phase.[36]
Muscle activation. All EMG signals were high-pass filtered using a recursive, second-order Butterworth filter at 35 Hz to remove any unwanted low-frequency movement artifact. Subsequently, EMG signals were visually inspected to identify any remaining artifact, which was defined as an abnormal spike in the signal, typically associated with foot contact. Any remaining artifacts resulted in the EMG data for that particular stride being excluded from further analysis. After DC-offset removal, root mean square (RMS) signal amplitude was calculated across all EMG data using a moving window of 50 ms to generate an EMG envelope. The EMG envelope for each muscle was then normalized to the maximum RMS amplitude found across all conditions for the respective muscle. The normalized peak EMG RMS envelope amplitude was calculated during stance phase; in addition, the normalized mean EMG RMS was calculated during both the stance and swing phases, allowing for comparisons in magnitude of both stance and swing phase muscle activation between HFS and FFS conditions.
Statistics. For each individual, the motion, force and EMG data were time normalized and ensemble averaged across a minimum of eight stride cycles to form individual variable means for each condition. Paired Student's t-tests were used to describe the influence of foot-strike pattern on stride temporal characteristics, peak GRF, joint kinematics and kinetics, and muscle activation. Statistical differences were established at P ≤ 0.05. Effect sizes (ES) are presented as standardized mean differences (Cohen's D), with ≤0.70 representing a small effect, >0.70 and <1.2 representing a moderate effect, and ≥1.2 representing a large effect.[37] Results are presented as mean ± SD unless otherwise stated.
Results
Stride Temporal Characteristics
Participants ran with similar stride durations (ES = 0.07, P = 0.54) and ground contact times (ES = 0, P = 0.77; Table 1), regardless of the orientation of their ankles at ground contact.
Ground Reaction Forces
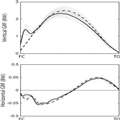
When running with an HFS pattern, vertical GRF–time profiles displayed a characteristic initial early impact transient, followed by a second, much later, larger peak (Figure 1). The initial impact peak transient seen for HFS running was absent when the participants ran with a forefoot technique (Figure 1); however, the magnitude of the peak vertical GRF was significantly higher with the forefoot technique (ES = 0.81, P ≤ 0.05; Table 1). Propulsive and breaking force was similar between conditions (Figure 1).
Figure 1.
Group mean ± SD (shaded area) for vertical GRF (top) and anterior–posterior GRF (bottom). Data are recorded from each participant running with a habitual (solid) and forefoot running technique (dashed) at 2.8 ms−1. Data are shown for stance only, from right foot contact (FC) to right toe-off (TO). All data are normalized to BW. *denotes a significant difference in peak vertical GRF between the running conditions.
Joint Kinematics
When running with an HFS pattern, participants generally made initial ground contact with the heel or midfoot. One participant landed with the ankle in slight plantar flexion (5.4°), whereas the remaining participants landed with the ankle in slight dorsiflexion (range, −8.5° to −2.0°; Figure 2). None of the participants landed with an FFS as their HFS pattern. Participants landed with their ankles in a significantly more plantar flexed orientation when instructed to run with an FFS (range, 6.4°–20.6°; ES = 3.5; P≤ 0.05, Table 2). Peak ankle dorsiflexion occurred in midstance for both conditions and was significantly greater when running with an HFS (ES = 0.88, P ≤ 0.05; Table 2). At toe-off, the ankle was in a more plantar flexed orientation in the forefoot running condition (ES = 1.1, P ≤ 0.05; Table 2).

Figure 2.
Group mean ± SD (shaded area) for joint angle (top), normalized joint moment (middle), and normalized joint power (bottom) calculated during stance phase for the midfoot (left), rearfoot (middle), and ankle (right). Data are recorded from each participant running with a habitual (solid) and forefoot running technique (dashed) at 2.8 ms−1. Data are shown for stance only, from right foot contact (FC) to right toe-off (TO). Vertical dashed lines denote the time point that the COP progressed anterior to the axis of rotation of the midfoot joint in the HFS condition. Joint work is calculated by trapezoidal integration of the joint power data. δ denotes a significant difference between running conditions in joint angle (at FC, peak, or TO) for each joint. γ denotes significant difference in peak joint moment between the running conditions. β denotes a significant difference in peak joint power (negative or positive), and α denotes a significant difference in negative or positive joint work between running conditions.
The rearfoot rotated in a similar manner to the ankle, with dorsiflexion occurring from forefoot contact through to midstance and plantar flexion occurring from midstance to toe-off. The rearfoot was in a more plantar flexed orientation at foot contact when running with an FFS (ES = 1.1, P ≤ 0.05; Table 2 and Figure 2). Peak rearfoot dorsiflexion occurred in midstance and was reduced in magnitude when running with an FFS pattern (ES = 3.5, P ≤ 0.05; Table 2). The rearfoot was more plantar flexed at toe-off with the forefoot running technique (ES = 1.1, P ≤ 0.05; Table 2).
For both running conditions, the midfoot dorsiflexed (LA compression) from early stance through to midstance (Figure 2) and subsequently plantar flexed through to toe-off. At foot contact, the midfoot joint was more plantar flexed when running with an FFS (ES = 2.7, P ≤ 0.05; Table 2), indicating a higher LA at foot contact. However, there was no difference in peak midfoot dorsiflexion between running conditions (ES = 0.06, P = 0.63; Table 2). The midfoot was in a position of greater plantar flexion at toe-off when participants ran with an FFS (ES = 0.9, P ≤ 0.05; Table 2).
Joint Kinetics
Peak ankle plantar flexion moment was greater when running with an FFS pattern (ES = 1.3, P ≤ 0.05; Table 2 and Figure 2). Peak negative ankle power (ES = 2.4, P ≤ 0.05; Table 2) and positive ankle power (ES = 0.74, P ≤ 0.05; Table 2) were also greater when running with an FFS technique. A greater amount of negative work (ES = 2.1, P ≤ 0.05; Table 2) and positive work (ES = 1.0, P ≤ 0.05, respectively) was performed at the ankle when running with an FFS.
For both running conditions, peak rearfoot plantar flexion moment occurred in midstance (Figure 2). Peak rearfoot plantar flexion moment was greater when running with a forefoot technique (ES = 1.3, P ≤ 0.05; Table 2 and Figure 2). Peak negative power was greater when running with a forefoot technique (ES = 1.6, P ≤ 0.05; Table 2); however, there was no difference in peak positive power between running conditions (ES = 0.3, P = 0.13; Table 2). A greater amount of negative work (ES = 1.8, P ≤ 0.05; Table 2) and positive work (ES = 0.6, P ≤ 0.05; Table 2) was performed at the rearfoot in the forefoot running condition.
In a similar manner to the ankle and rearfoot, peak midfoot plantar flexion moment occurred in midstance for both running conditions (Figure 2). Peak midfoot plantar flexion moment was greater when running with a forefoot technique (ES = 1.1, P ≤ 0.05; Table 2 and Figure 2). Peak negative midfoot power (ES = 2.7, P ≤ 0.05; Table 2) and positive midfoot power (ES = 0.77, P ≤ 0.05; Table 2) were also greater when running with an FFS. A greater amount of negative work (ES = 2.2, P ≤ 0.05) and positive work (ES = 1.1, P ≤ 0.05; Table 2) was also performed at the midfoot when running with a forefoot technique, suggesting that a greater magnitude of mechanical energy is absorbed, returned, and/or generated about the midfoot. Despite similar peak deformation and an increased plantar flexion moment, quasi-stiffness of the midfoot reduced when running with a forefoot technique (ES = 0.65, P ≤ 0.05; Table 2).
Muscle Activation
The AH muscle displayed a large burst of activity that commenced during late swing phase and continued through to toe-off, with peak activation occurring in midstance (Figure 3). When running with an FFS, mean AH activity was greater in swing (ES = 0.67, P ≤ 0.05; Table 3) and stance (ES = 0.63, P ≤ 0.05; Table 3) phases. Peak stance-phase muscle activation was also greater when running with an FFS (ES = 0.71, P ≤ 0.05; Table 3).

Figure 3.
Group mean ensembles ± SD (shaded area) for EMG normalized RMS signal amplitude for MG, SOL, FDB and AH. Group mean ensembles are defined fromtoe-off (TO) to ipsilateral toe-off for the right foot. Data recorded during running at 2.8 ms−1. For each muscle EMG data is normalized to the maximal amplitude recorded for all trials. The HFS condition is the solid lines and the forefoot condition the dashed lines. Vertical lines denote onset of stance phase at foot contact (FC). *denotes significant difference between running conditions.
The FDB activation pattern was characterized by a large burst of activity commencing at foot contact, with a midstance peak and deactivation shortly after toe-off (Figure 3). Because this muscle is primarily active in stance phase, mean swing-phase activation was similar between conditions (ES = 0.01, P = 0.95; Table 3). Mean FDB stance-phase activation substantially higher when running with an FFS (ES = 0.56, P ≤ 0.05; Table 3); however, there was no significant difference in peak stance-phase activation for this muscle (ES = 0.49, P = 0.19; Table 3).
For both conditions, MG displayed a large burst of activation commencing in late swing, continuing through stance, with deactivation occurring during late stance phase (Figure 3). Mean swing-phase activation was substantially greater when running with an FFS (ES = 2.36, P ≤ 0.05; Table 3), whereas mean (ES = 0.57, P = 0.09; Table 3) and peak (ES = 0.06, P = 0.84; Table 3) stance-phase activity was similar for both running conditions.
The SOL activation pattern was similar for both running conditions, with a single burst of activity that commenced in terminal swing phase, peaked during midstance, and ceased before toe-off (Figure 3). Swing-phase activation was comparable for both running techniques (ES = 0.2, P = 0.39), as was mean (ES = 0.50, P = 0.19) and peak (ES = 0.69, P = 0.21) stance-phase activity (Table 3).
Discussion
This study provides novel evidence for active alterations to the mechanical function of the foot when acutely switching between non–forefoot and forefoot running techniques. When running at matched velocities, forefoot running resulted in higher midfoot joint moments, more positive and more negative work being performed about the midfoot. Increases in joint moments for forefoot running occurred concurrently with increased intrinsic foot muscle activation and no increase in peak midfoot dorsiflexion. Thus, it seems that the intrinsic foot muscles may be activating to increase energy absorption and return, while also preventing excessive deformation of the midfoot when acutely transitioning to a forefoot technique.
LA Kinematics
Peak LA deformation was not influenced by foot-strike technique, despite the increased joint moments about the midfoot when running on the forefoot. This finding is analogous to a number of previous studies that also reported no difference in peak LA deformation when running with rearfoot or forefoot running techniques.[4,14,16] Interestingly, the previous studies by Wager and Challis[14] and McDonald et al.[16] have made the assumption in their modeling that the foot is a passive structure with no muscular control of LA stiffness. The findings of our current study highlight that the foot is not passive and the plantar aponeurosis is not the sole structure responsible for maintaining LA stiffness during running. If the LA compression was indeed controlled purely by passive elastic structures (e.g., plantar aponeurosis), the increased torques observed when running with a forefoot technique would produce an increase in LA deformation.[12,38] This was not the case in the current study or in the previously mentioned studies,[4,14,16] indicating that active muscular control must be contributing to control of LA mechanics when switching between foot-strike techniques.
Recently published studies performed within our laboratory have highlighted the importance of the plantar intrinsic foot muscles in actively controlling the LA during quiet stance and locomotion.[15,24,26] When activated, these muscles have the capacity to provide buttressing support for the LA, resisting excessive deformation that may occur at loads exceeding BW.[23]Furthermore, it seems that the CNS can vary the relative contribution of these muscles in controlling the LA, depending on the magnitude of forces encountered when running.[15] On the basis of our knowledge of the function of the intrinsic foot muscles,[15,23] it becomes apparent that the observed increase in AH and FDB activation during stance may be responsible for the lack of difference in peak LA deformation between running techniques, despite obvious differences in joint moments. An increase in force output from the intrinsic foot muscles when running with a forefoot technique may serve to reinforce the LA in midstance, preventing excessive LA deformation and any associated increase in plantar aponeurosis strain. Recently, McDonald et al.[16] suggested that the risk of injury to the plantar aponeurosis (i.e., plantar fasciopathy) might not be influenced by foot-strike technique, owing to the similarity in peak plantar aponeurosis strains between techniques. The current study reveals that there is a substantial increase in loading and energy absorbed about the midfoot when running with a forefoot technique and that this increase is likely associated with an increase in force output from the plantar intrinsic foot muscles. Therefore, without specific training, habitual rearfoot runners may not have the strength or endurance in the intrinsic foot muscles to sustain a forefoot running technique for prolonged periods. This may subsequently influence the loading on the plantar aponeurosis. Further investigations may now be warranted to determine the time required to sufficiently strengthen the intrinsic foot muscles, to safely transition to a forefoot running technique.
We quantified midfoot quasi-stiffness by dividing the change in joint moment by the change in joint angle during the first half of stance.[36] Using this approach, we observed a reduction in quasi-midfoot stiffness when participants ran with a forefoot technique, despite increased peak midfoot moments and similar peak midfoot deformation. This finding is similar to a number of previous investigations.[4,14,16] However, it may seem counterintuitive, because the lack of change in peak LA deformation and the increase in the associated peak moment could also imply an increase in quasi-midfoot stiffness. This highlights the complexity of interpreting quasi-stiffness about joints during dynamic tasks.[39] More importantly, the data emphasize that the observed joint mechanics cannot be explained by passive elastic structures like the plantar fascia alone, because the mechanics are inconsistent with those of a simple elastic spring.[12] In the forefoot running condition, AH activation increased substantially in late swing, preceding an increase in midfoot plantar flexion and LA height before foot contact. This alteration in late swing mechanics allowed the midfoot to move through a greater total range of motion during stance phase, reducing the quasi-stiffness of the midfoot without altering peak midfoot deformation and, by association, peak plantar fascia strain.[16] Therefore, the increased loading within the midfoot must be shared with other tissues, such as the intrinsic foot muscles. Perhaps the observed increase in activation of intrinsic foot muscles during forefoot running helps to limit strain of the fascia, reducing the risk of strain reaching harmful levels.
If strain within the plantar fascia does not increase in forefoot running, there are implications for how we interpret the increased energy storage and return in this condition. The increased energy storage about the LA in forefoot running was related to a greater range of motion about the midfoot and an increase in midfoot moments. Given that peak strain primarily determines the magnitude of stored energy in passive elastic structures,[13] it is apparent that the plantar fascia is not responsible for the increased absorption of energy in the forefoot running condition. This indirect evidence, in conjunction with the observed greater intrinsic muscle activation in forefoot running, implies strongly that the intrinsic foot muscles are contributing to the increased energy absorption about the LA in forefoot running. This suggestion is supported by the fact that negative work at the LA (energy absorption) increased by 240%, but the subsequent positive work (including energy returned) increased by just 42%. If energy were being absorbed only by elastic structures, we would expect a much greater proportion of the energy absorbed to be returned. Therefore, it is likely that any increase in mechanical work performed about the LA when running with a forefoot technique occurs because of active muscle contraction and may incur an increased metabolic cost. An unknown in this interpretation is how much energy could be stored and returned in series elastic tissues of the muscles themselves.
Joint Moments
Forefoot running resulted in higher plantar flexion moments being generated about the ankle and midfoot. When running with a forefoot technique, the resultant GRF is generally positioned within the anterior portion of the foot, as the forefoot makes initial contact with the ground. Conversely, with a rearfoot technique, the COP is initially located on the lateral aspect of the heel and progresses forward as the body progresses over the foot.[2,17,40] If the COP is located further anterior when running with a forefoot technique, it will have a greater moment arm about the ankle and midfoot, which, alongside an increase in peak vertical GRF, likely explains the increase in plantar flexion moments that are generated about these joints. Although the finding of an increase in plantar flexion moment about the midfoot is quite novel, a similar increase in ankle joint moment has been reported in a number of previous studies examining differences in ankle joint kinetics when switching between rearfoot and forefoot running techniques.[19,41]
Mechanical Work and Power
The energetic benefits of the springlike function of the LA during running have been well documented, with this mechanism providing between 8% and 17% of the mechanical energy required for each stride via the elastic stretch and recoil of the plantar aponeurosis.[12,13] Previous studies exploring the influence of foot-strike technique on elastic energy storage in the LA[14,16] have reported that a forefoot running technique does not promote increased utilization of the elastic energy, because of the lack of difference in peak plantar aponeurosis strain between running techniques. Within our current experiment, we observed an increase in mechanical power absorption and generation and an associated increase in negative and positive work about the LA, in the absence of any additional increase in LA deformation. The plantar intrinsic foot muscles are known to actively lengthen and shorten during the stance phase of running and therefore have the potential to absorb and generate mechanical power about the LA.[15] The increase in intrinsic foot muscle activation when running with a forefoot technique indicates that the contribution of these muscles to mechanical work about the LA increases when running with a forefoot technique. The intrinsic foot muscles have relatively long tendons and short muscle fibers[20] and thus may be well suited to elastic energy storage and return. The additional negative work done at the midfoot joint when running with a forefoot technique may allow for greater utilization of the foot-spring mechanism, allowing additional energy to be stored in the tendons of the intrinsic foot muscles, without the potentially detrimental effects of excessive strain on the plantar aponeurosis. In this sense, we might consider the intrinsic foot muscles as reserve actuators, able to act in parallel with the plantar aponeurosis when a certain level of force generation is required. However, it must also be noted that the increase in work performed about the LA when running with a forefoot technique may not translate directly into metabolic energy savings, owing to the added energetic cost of muscle activation required to perform this work. The increased cost of activation in the intrinsic foot muscles and also the posterior leg muscles when running with a forefoot technique may in part explain the lack of difference in energetic cost between habitual rearfoot and forefoot runners[42,43]because the increased utilization of springlike running mechanics may be offset by the additional energetic cost of muscle activation.
Runners with an habitual forefoot technique generally display shorter ground contact times, shorter stride length, and an increased cadence compared with runners who are habitual rearfoot strikers.[3,19,43] In the current experiment, our group of habitual rearfoot runners maintained relatively constant stride temporal characteristics when acutely changing to a forefoot technique. This finding is in line with a number of previous studies that have shown a lack of immediate change in stride temporal characteristics as a result of an acutely imposed forefoot technique.[44] Thus, it is apparent that changes in stride temporal characteristics do not occur automatically when changing foot-strike technique. Specific training to encourage a shorter stride length and increased cadence may also need to be included as part of a habituation process in a transition to forefoot running.
Methodological Considerations
The LA is a multiarticular structure.[28] We have modeled this structure as single functional midfoot joint. This technique allows for important general insights into the function of the LA; however, it may also be an oversimplification of the complex sequence of rotations and translations that occur about a series of small joints within this overall structure. Furthermore, we have only modeled the function of this structure in the sagittal plane. It is likely that motion of the LA also occurs in the frontal and transverse planes.[23,25]
The use of an inverse dynamic solution allowed us to provide unique insight into the mechanical function of the LA when running with rearfoot and forefoot running techniques. A limitation of this approach is that it is difficult to quantify the relative percentage of GRF that is acting on the forefoot segment at any given time in stance, because some force may be simultaneously applied to the rearfoot. However, because the COP rapidly progresses anteriorly in stance phase and the forefoot bears a substantial portion of the GRF force for the majority of stance duration,[2,17] any errors associated with this assumption are likely to be small and localized to the initial period of stance in the habitual running condition. To mitigate this limitation, we have calculated rearfoot and midfoot moments and powers after the COP has progressed anterior to the axis of the midfoot joint. The COP passed the axis of the midfoot at approximately 25% stance phase in the habitual running condition, and after this time, we are confident that the vast majority of contact forces are applied to the forefoot. When running with a forefoot running technique, the COP was anterior to the axis of the midfoot joint for the entire duration of stance phase; therefore, the GRF is applied to the forefoot segment from the instant of foot contact. If small errors do exist on the basis of our GRF assignment assumption, these are limited to the rearfoot running condition and will lead to an overestimation in LA joint moments; therefore, the magnitude of differences in joint kinetics between conditions may actually be slightly larger than those reported. Providing further support to the validity of our inverse dynamic approach, the joint moments at the rearfoot are comparable to the ankle joint moments using a traditional rigid foot segment, both within our current study and also to the values presented in previous studies reporting ankle joint moments during running at similar speeds.[45]
Our definition of quasi-midfoot stiffness assumes that the force deformation properties of the midfoot are linear. Acting in parallel with the plantar fascia, the intrinsic muscles have the capacity to modify the stiffness of the midfoot via CNS control;[23] therefore, it is likely that this structure may display nonlinear force deformation characteristics. With this in mind, it is likely that the measure of quasi-joint stiffness applied in this study may not adequately reflect complex nonlinear behavior of the midfoot.[39]
Participants within this experiment were shod rearfoot runners who were instructed to run with their HFS technique and a forefoot technique. It is possible that different observations may have been made if habitual forefoot runners were included in this experiment. However, on the basis of the findings of previous studies,[13,16] it is unlikely that LA kinematics would differ between habitual rearfoot and forefoot runners when instructed to run with forefoot and rearfoot techniques. Furthermore, given that activation of the intrinsic foot muscles is dependent on loading requirement,[23] it is unlikely that the activation patterns of these muscles would substantially differ either.
Conclusions
Foot-strike technique substantially influences the mechanical function of the LA during running. A forefoot running technique leads to an increase in loading about the midfoot and an increase in positive and negative mechanical work performed by the intrinsic foot muscles. These findings suggest that adequate strength and endurance training for the intrinsic foot muscles should be included as part of training programs for runners seeking to transition from a rearfoot to a forefoot running technique.
- Altman AR, Davis IS. A kinematic method for footstrike pattern detection in barefoot and shod runners. Gait Posture. 2012;35(2):298–300.
- Cavanagh PR, Lafortune MA. Ground reaction forces in distance running. J Biomech. 1980;13(5):397–406.
- Lieberman DE, Venkadesan M, Werbel WA, et al. Foot strike patterns and collision forces in habitually barefoot versus shod runners. Nature. 2010;463(7280):531–5.
- Perl DP, Daoud AI, Lieberman DE. Effects of footwear and strike type on running economy. Med Sci Sports Exerc. 2012;44(7):1335–43.
- Altman AR, Davis IS. Barefoot running: biomechanics and implications for running injuries. Curr Sports Med Rep. 2012;11(5):244–50.
- Hasegawa H, Yamauchi T. Foot strike patterns of runners at the 15-km point during an elite-level half marathon. J Strength Cond Res. 2007;21(3):888–93.
- Larson P, Higgins E, Kaminski J, et al. Foot strike patterns of recreational and sub-elite runners in a long-distance road race. J Sports Sci. 2011;29(15):1665–73.
- Daoud AI, Geissler GJ, Wang F, Saretsky J, Daoud YA, Lieberman DE. Foot strike and injury rates in endurance runners: a retrospective study. Med Sci Sports Exerc. 2012;44(7):1325–34.
- Lieberman DE, Bramble DM. The evolution of marathon running: capabilities in humans. Sports Med. 2007;37(4–5):288–90.
- Diebal AR, Gregory R, Alitz C, Gerber JP. Forefoot running improves pain and disability associated with chronic exertional compartment syndrome. Am J Sports Med. 2012;40(5):1060–7.
- Heiderscheit BC. Gait retraining for runners: in search of the ideal. J Orthop Sports Phys Ther. 2011;41(12):909–10.
- Ker RF, Bennett MB, Bibby SR, Kester RC, Alexander RM. The spring in the arch of the human foot. Nature. 1987;325:147–9.
- Stearne SM, McDonald KA, Alderson JA, North I, Oxnard CE, Rubenson J. The foot's arch and the energetics of human locomotion. Sci Rep. 2016;6:19403.
- Wager JC, Challis JH. Elastic energy within the human plantar aponeurosis contributes to arch shortening during the push-off phase of running. J Biomech. 2016;49(5):704–709.
- Kelly LA, Lichtwark G, Cresswell AG. Active regulation of longitudinal arch compression and recoil during walking and running. J R Soc Interface. 2015;12(102):20141076.
- McDonald KA, Stearne SM, Alderson JA, North I, Pires NJ, Rubenson J. The role of arch compression and metatarsophalangeal joint dynamics in modulating plantar fascia strain in running. PLoS One. 2016;11(4):e0152602.
- Cavanagh PR. A technique for averaging center of pressure paths from a force platform. J Biomech. 1978;11(10–12):487–91.
- Nilsson J, Thorstensson A. Ground reaction forces at different speeds of human walking and running. Acta Physiol Scand. 1989;136(2):217–27.
- Stearne SM, Alderson JA, Green BA, Donnelly CJ, Rubenson J. Joint kinetics in rearfoot versus forefoot running: implications of switching technique. Med Sci Sports Exerc. 2014;46(8):1578–87.
- Kura H, Luo ZP, Kitaoka HB, An KN. Quantitative analysis of the intrinsic muscles of the foot. Anat Rec. 1997;249(1):143–51.
- Ledoux WR, Hirsch BE, Church T, Caunin M. Pennation angles of the intrinsic muscles of the foot. J Biomech. 2001;34(3):399–403.
- Tosovic D, Ghebremedhin E, Glen C, Gorelick M, Brown JM. The architecture and contraction time of intrinsic foot muscles. J Electromyogr Kinesiol. 2012;22(6):930–8.
- Kelly LA, Cresswell AG, Racinais S, Whiteley R, Lichtwark G. Intrinsic foot muscles have the capacity to control deformation of the longitudinal arch. J R Soc Interface. 2014;11(93):20131188–8.
- Kelly LA, Lichtwark GA, Farris DJ, Cresswell A. Shoes alter the spring-like function of the human foot during running. J R Soc Interface. 2016;13(119). doi: 10.1098/rsif.2016.0174.
- Leardini A, Benedetti MG, Berti L, Bettinelli D, Nativo R, Giannini S. Rear-foot, mid-foot and fore-foot motion during the stance phase of gait. Gait Posture. 2007;25(3):453–62.
- Kelly LA, Kuitunen S, Racinais S, Cresswell AG. Recruitment of the plantar intrinsic foot muscles with increasing postural demand. Clin Biomech (Bristol, Avon). 2012;27(1):46–51.
- Kelly LA, Racinais S, Cresswell AG. Discharge properties of abductor hallucis before, during, and after an isometric fatigue task. J Neurophysiol. 2013;110(4):891–8.
- Hicks JH. The mechanics of the foot. I. The joints. J Anat. 1953;87:345–57.
- Hicks JH. The mechanics of the foot. II. The plantar aponeurosis and the arch. J Anat. 1954;88:25–30.
- Caravaggi P, Pataky T, Günther M, Savage R, Crompton R. Dynamics of longitudinal arch support in relation to walking speed: contribution of the plantar aponeurosis. J Anat. 2010;217(3):254–61.
- Dixon PC, Böhm H, Döderlein L. Ankle and midfoot kinetics during normal gait: a multi-segment approach. J Biomech. 2012;45(6):1011–6.
- Winter DA. Energy generation and absorption at the ankle and knee during fast, natural, and slow cadences. Clin Orthop Relat Res. 1983;(175):147–54.
- De Leva P. Adjustments to Zatsiorsky–Seluyanov's segment inertia parameters. J Biomech. 1996;29(9):1223–30.
- Stefanyshyn DJ, Nigg BM. Contribution of the lower extremity joints to mechanical energy in running vertical jumps and running long jumps. J Sports Sci. 1998;16(2):177–86.
- Edwards WB, Troy KL, Derrick TR. On the filtering of intersegmental loads during running. Gait Posture. 2011;34(3):435.
- Farley CT, Morgenroth DC. Leg stiffness primarily depends on ankle stiffness during human hopping. J Biomech. 1999;32(3):267–73.
- Hopkins W. A New View of Statistics. 2017:1–26 [cited 2017 July]. Available from: http://wwwsportsciorg/resource/stats.
- Erdemir A, Hamel AJ, Fauth AR, Piazza SJ, Sharkey NA. Dynamic loading of the plantar aponeurosis in walking. J Bone Joint Surg Am. 2004;86-A(3):546–52.
- Latash ML, Zatsiorsky VM. Joint stiffness: myth or reality? Hum Mov Sci. 1993;12(6):653.
- Novacheck TF. The biomechanics of running. Gait Posture. 1998;7:77–95.
- Kulamala J, Avela J, Pasanen K, Parkkari J. Forefoot strikers exhibit lower running-induced knee loading than rearfoot strikers. Med Sci Sports Exerc. 2013;45(12):2306–13.
- Ardigò LP, Lafortuna C, Minetti AE. Metabolic and mechanical aspects of foot landing type, forefoot and rearfoot strike, in human running. Acta Physiol Scand. 1995;155:17–22.
- Gruber AH, Umberger BR, Braun B, Hamill J. Economy and rate of carbohydrate oxidation during running with rearfoot and forefoot strike patterns. J Appl Physiol (1985). 2013;115(2):194–201.
- Almeida MO, Davis IS, Lopes AD. Biomechanical differences of foot-strike patterns during running: a systematic review with meta-analysis. J Orthop Sports Phys Ther. 2015;45(10):738–55.
- Schache AG, Blanch PD, Dorn TW, Pandy MG, Rosemond D, Brown NA. Effect of running speed on lower limb joint kinetics. Med Sci Sports Exerc. 2011;43(7):1260–71.